Ce billet écrit par des étudiantes et étudiants de Master 1 du Magistère Européen de Génétique est co-publié sur le site de nos collaborateurs pédagogiques – Papier-mâché Promotion 2023-2024
Écriture : Elise Mary, François Bouvard, Gabrielle Maillard, Paloma Guernalec Relecture scientifique : Équipe pédagogique : Sandra Claret et Alain Zider Relecture de forme : Lucile Rey Temps de lecture : environ 12 minutes. Thématiques : Biologie cellulaire, Neurobiologie (Biologie) Publication originale : Brault J.-B., et al., RAB6 and dynein drive post-Golgi apical transport to prevent neuronal progenitor delamination. EMBO Reports, 2022. DOI : 10.15252/embr.202254605 → Des notions POUR APPROFONDIR à la fin de l’article.

On se demande tous comment le cerveau parvient à être aussi complexe, mais auriez-vous pensé que son développement repose en partie sur le transport d’une petite molécule ? En suivant l’article du Dr. Brault et de ses collaborateurs, vous allez vous plonger dans le trajet quotidien de Crumbs, une molécule particulière : une protéine impliquée dans l’intégrité du tissu neural. Quel est son rôle ? Quel est son trajet ? Et que se passerait-il si elle loupait son train ?
Une caméra dans les cellules
Et si le moindre problème provoquant l’arrêt d’une seule ligne de train chamboulait toute la France ? C’est un peu ce qu’il se passe dans les cellules d’un cerveau en cours de développement lorsque le transport de la protéine Crumbs est affecté.
Le cerveau est l’organe le plus complexe du corps chez les mammifères, mais aussi le moins connu. Grâce à la technologie d’imagerie subcellulaire en direct [voir Pour approfondir], les chercheurs de l’équipe du Dr. Baffet ont filmé ce qu’il se passe dans les cellules qui composent le cerveau de souris ! C’est une véritable révolution pour mieux comprendre son développement, et nous allons justement utiliser cette technologie pour voyager ensemble au cœur du cerveau et suivre en direct le transport des protéines dans les cellules neurales. Tout comme l’équipe du Dr. Baffet, nous avons organisé ce voyage autour d’une question centrale : en quoi ce système de transport de molécules Crumbs vers la membrane des cellules est-il si essentiel au développement du cerveau ?
Tout va bon train
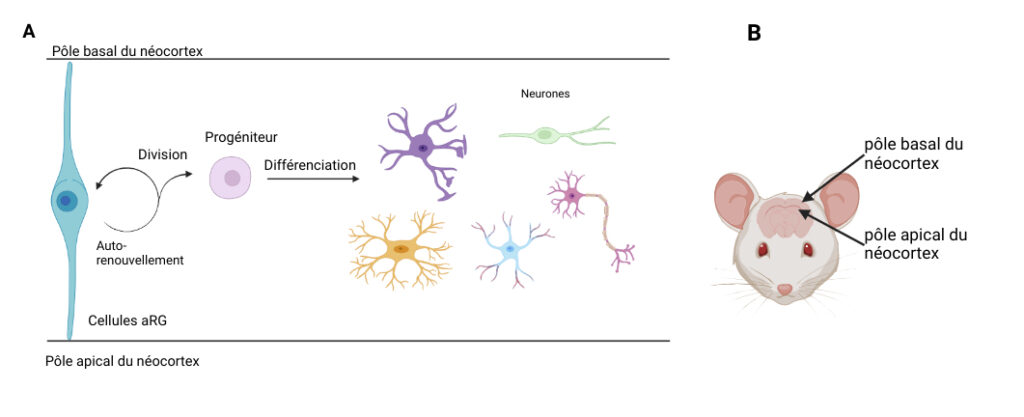
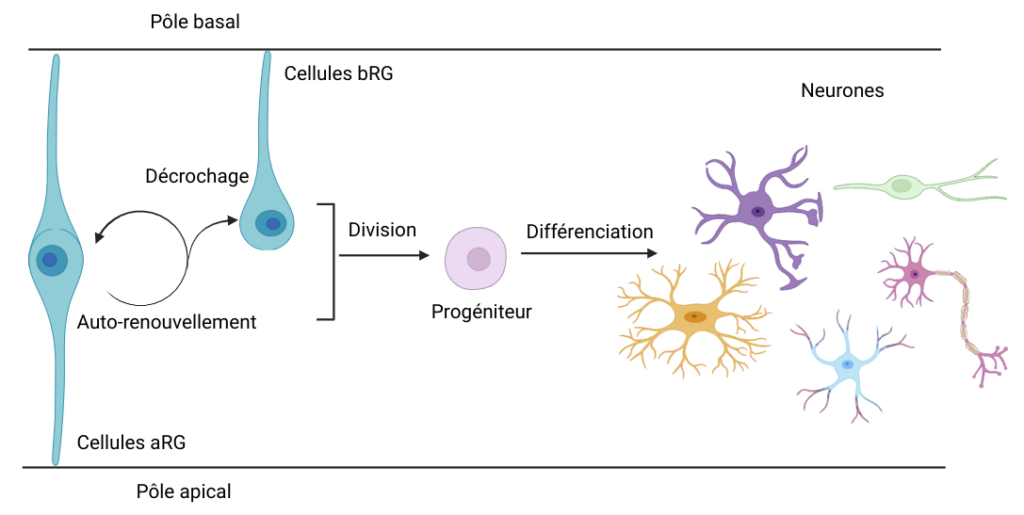
Entrons tout d’abord dans la boîte crânienne de la souris. Étonnant ! Son cerveau est tout lisse. Contrairement à celui de l’espèce humaine, il ne possède pas de replis ! Le cerveau est composé de plusieurs parties, dont le néocortex, qui nous intéresse ici. Présent uniquement chez les mammifères [1], il constitue la couche externe du cerveau. Il peut se replier … ou pas, comme chez la souris. D’ailleurs, saviez-vous que le néocortex possède des pôles ? Pas le pôle Nord ni le pôle Sud, mais un pôle apical, situé vers l’intérieur du néocortex, et un pôle basal, vers l’extérieur [Figure 1]. Quoiqu’il en soit, avant qu’il soit lisse ou plié, notre cerveau n’est qu’un petit amas de cellules.
Approchons-nous un peu plus de ce néocortex en cours de développement, plus exactement au stade embryonnaire. On peut y apercevoir des cellules particulièrement intéressantes : les cellules souches neurales. Ces cellules sont très importantes car elles permettent de générer, par multiplication, toutes les cellules composant notre cerveau, dont les neurones ! Elles se distinguent en deux types de neurones progéniteurs [*] : les cellules aRG (cellules apicales de la Glie Radiaire), accrochées à la fois au pôle apical et basal du néocortex [Figure 1], et les cellules bRG (cellules basales de la Glie Radiaire), accrochées seulement au pôle basal. Les souris possèdent en grande majorité des cellules aRG [2], alors que l’espèce humaine, elle, possède les deux types de cellules en quantité équivalente.
Continuons notre exploration, et dirigeons-nous maintenant vers l’intérieur des cellules aRG. On y trouve des microtubules, des structures qui ressemblent à de grands rails qui les traversent. Les microtubules sont des constituants du squelette de la cellule. Ils sont chargés du transport de différentes protéines via les trains (il en existe plusieurs dans la cellule : vésicules [**], complexes protéiques, etc.) qui circulent dessus. En continuant notre exploration, on découvre la grande gare de la cellule, de laquelle partent de nombreuses protéines : l’appareil de Golgi. L’appareil de Golgi est essentiel pour contrôler le tri et l’envoi des protéines vers des compartiments spécifiques de la cellule (comme le noyau), ou vers la membrane plasmique, une barrière qui délimite les cellules. Cette gare se compose de deux parties : une partie où les protéines finissent de se préparer (on appelle cela la maturation), et une autre partie où les protéines attendent sur les quais avant de monter dans leur train pour être amenées à leur destination et exercer leur rôle.
Une protéine, RAB6, est très abondante dans la gare. Elle est cruciale pour réguler le transport cellulaire d’autres protéines par l’intermédiaire de vésicules. RAB6 se comporte donc un peu comme un chef de bord qui gère les départs des trains et le transport des voyageurs.
« Tutututu… Mesdames, messieurs, le train DYN à destination du pôle apical cellulaire est sur le point de partir. »
Vite, vite, la protéine Crumbs se dirige vers le quai pour aller travailler. Comme tous les matins, elle doit prendre le train pour se rendre au pôle apical des cellules aRG. Eh oui, certaines cellules aussi possèdent des pôles ! La protéine Crumbs, accompagnée de RAB6, rentre donc dans une vésicule (un petit wagon) tirée par la dynéine, un moteur moléculaire (la locomotive). Ainsi, le complexe vésicule-dynéine forme les trains qui circulent sur les microtubules, la dynéine tirant les vésicules jusqu’au terminus du train, qui se trouve ici au pôle apical de la cellule [Figure 2].
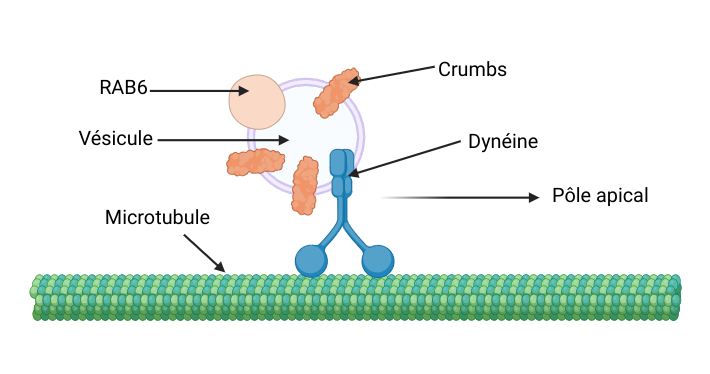
Crumbs peut donc effectuer sa journée de travail au pôle apical de la membrane plasmique. C’est une protéine transmembranaire, c’est-à-dire qu’elle traverse la membrane de la cellule de chaque côté. Son rôle principal est d’accrocher les cellules aRG à la surface apicale du néocortex grâce à des jonctions adhérentes. Ces jonctions agissent comme une sorte de « colle » entre les cellules. Elles assurent donc la cohésion et le maintien des différentes structures cérébrales entre elles. L’accrochage des cellules aRG au pôle apical permet aux couches de tissu neural, ou neuroépithélium, de se former correctement. Le néocortex peut ainsi se développer.
Un retard pour cause de protéines sur les voies
« Tutututuuu… Voie 12, le train DYN à destination du pôle apical de la cellule est retardé en raison d’un obstacle sur la voie. »
Encore des problèmes de transport… Le train que Crumbs devait prendre est bloqué sur les voies à cause d’un obstacle, la dynarrestin, une protéine inhibitrice de la dynéine : son rôle est de l’empêcher d’avancer sur les microtubules. Mais… qui a donc placé cet obstacle ici ? Les responsables sont les chercheurs, qui l’ont utilisée pour tester l’impact de l’absence de transport sur le développement du néocortex. Mauvaise nouvelle : Crumbs ne sera pas à l’heure sur son lieu de travail, le pôle apical de la cellule.
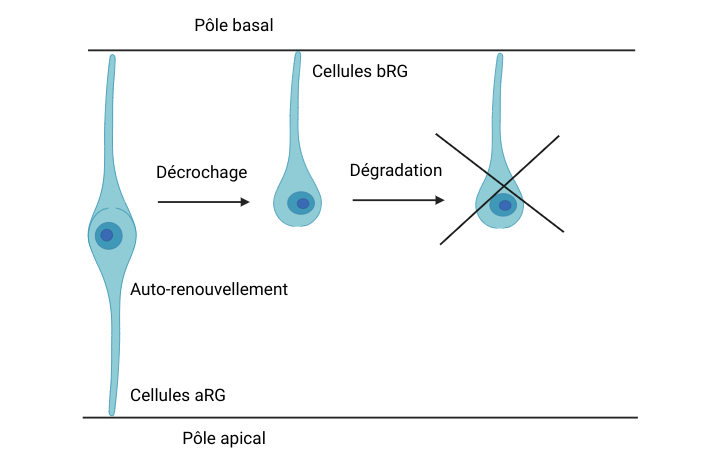
Une alerte apparaît soudainement sur les écrans de la gare : toutes les cellules aRG sont en train de se décrocher du neuroépithélium ! Crumbs est recherchée, où est-elle ? Le cerveau commence à rétrécir !
Son train ne pouvant pas partir, la protéine Crumbs reste sur le quai. Son absence entraîne le décrochage des cellules aRG du pôle apical du néocortex, un processus que l’on appelle la délamination. Les cellules aRG deviennent alors des cellules bRG, c’est-à-dire des cellules de la glie radiaire uniquement accrochées au pôle basal du néocortex.
Mais que fait le chef de gare ?
Le phénomène de délamination [Figure 3], provoqué par les problèmes de transport, est extrêmement rare chez les souris. Elles ont très peu de cellules bRG et le décrochage des aRG du pôle apical déclenche une désorganisation du neuroépithélium et une perturbation de son intégrité, ce qui peut mener à une microcéphalie, une diminution de la taille du cerveau. Les chercheurs ont émis l’hypothèse que la régulation du transport, notamment de Crumbs, est déterminante dans l’accrochage des cellules aRG chez les souris. Cette régulation, qui expliquerait la faible quantité de cellules bRG, reste pourtant largement inexplorée.
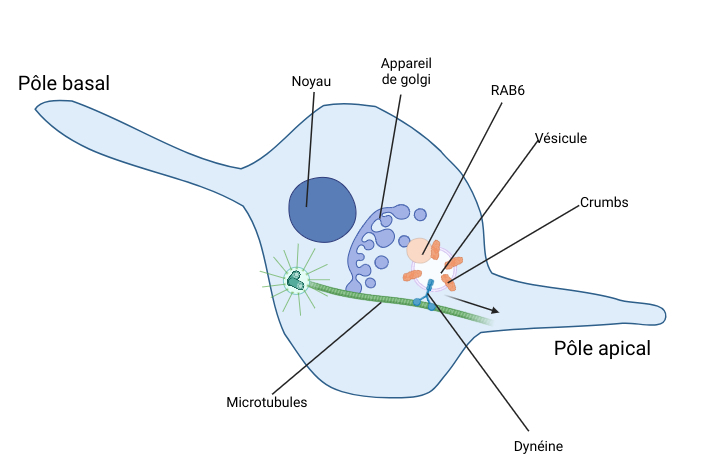
Les chercheurs se sont donc ensuite demandé si le même phénomène aurait pu être causé par une grève des chefs de bord, les protéines RAB6, et en particulier de tester son effet sur le transport de Crumbs. En effet, ces protéines permettent le départ et le transport des vésicules contenant des protéines comme Crumbs à la sortie de la gare Golgi [Figure 4]. Pour répondre à leur question, le Dr. Baffet et ses collaborateurs ont utilisé une autre méthode que celle utilisée avec la protéine Crumbs. Au lieu d’empêcher l’action de RAB6, ils ont décidé de l’enlever totalement des cellules en faisant ce qu’on appelle un knock-out. Autrement dit, ils ont empêché la construction de RAB6 en agissant directement au niveau de l’ADN. Lorsque les gènes RAB6 sont inactivés, les protéines qu’ils codent ne sont plus présentes et le transport ne se fait plus. Les trains sont coincés en gare. Crumbs ne peut donc pas non plus se rendre au travail.
Mais, attention ! Il existe en fait deux gènes RAB6 différents, codant pour deux protéines différentes : RAB6A et RAB6B. On a une redondance du rôle de RAB6A et RAB6B : quand l’une a un problème, l’autre la remplace et le travail est assuré ! Ainsi, l’inactivation d’une seule forme ne suffit pas à arrêter le transport. Les chercheurs ont donc procédé à un double knock-out, pour inactiver les deux gènes.
Sans les contrôleurs RAB6, Crumbs cherche à trouver une solution, elle ne veut pas que son absence cause un problème de développement du cerveau. D’un coup, elle voit qu’un train arrive sur le quai. Victoire ! Un train est en mesure de rouler ! Ni une ni deux, Crumbs monte à l’intérieur. Quelques minutes plus tard, le train se met en marche. Crumbs est sauvée ! Elle pense que le cerveau n’aura plus de problème.
« Tutututu… Le train KIN arrive en gare du pôle basal. La SNCF vous souhaite une bonne journée. »
Pôle basal ?!
Catastrophe ! Crumbs a emprunté un train kinésine et non un train dynéine ! La kinésine est comme une protéine dynéine mais elle va dans le sens opposé à cette dernière. En effet, lorsque le trafic de la locomotive dynéine est affecté, la locomotive kinésine prend le relais. Crumbs part alors dans le mauvais sens et se retrouve au pôle basal, à l’opposé de son lieu de travail. Elle ne pourra jamais retourner au pôle apical à temps, et la cellule va se décrocher…
Crumbs aurait dû arriver à l’heure
Ainsi, Crumbs doit accepter son sort, son problème de transport cause beaucoup de problèmes au niveau du néocortex. En effet, elle est indispensable au maintien de l’agencement de l’épithélium neural et son acheminement au pôle apical permet le contrôle de l’équilibre entre le maintien des cellules aRG et la délamination. Ce processus de délamination peut être problématique chez la souris et notamment causer une mauvaise orientation des pôles de la cellule.
Les résultats de l’article du Dr. Baffet et de ses collaborateurs donnent une piste pour comprendre pourquoi le cerveau des humains est plus complexe que celui des souris. En effet, chez les souris, la délamination des aRG en bRG n’est pas un processus naturel : lorsque le transport de Crumbs est affecté, on a un décrochage des aRG, qui ne peuvent plus s’auto-renouveler. Ceci induit une perte des cellules souches, qui peut expliquer le phénomène de microcéphalie. Au contraire, chez les êtres humains, ce processus de délamination est totalement normal, et même nécessaire au développement du néocortex [3]. Le cerveau de l’être humain est 20 fois plus gros que celui de la souris [4], et chez lui certaines cellules aRG deviennent spontanément des cellules bRG par décrochage du pôle apical [Figure 5].
Les chercheurs ont émis l’hypothèse que chez l’être humain, il existe une régulation, un contrôle du transport de Crumbs qui permet soit la multiplication des aRG, soit une formation de bRG.
Ainsi, le cerveau humain possèderait un système de régulation qui pourrait provoquer la formation d’un grand nombre de bRG, qui, lorsqu’elles se divisent, augmentent la taille du néocortex au pôle basal, ce qui expliquerait l’apparition de nombreuses circonvolutions. Les expériences présentées dans cet article ne permettent évidemment pas de savoir si ce système de régulation existe bel et bien chez l’espèce humaine. Cependant, il s’agit d’une première hypothèse qui ouvre la voie à de nombreuses expériences futures qui permettront de décortiquer les mystères du cerveau humain.
Éléments pour approfondir
Voyage au cœur des neurones
Lors d’une interview, le Dr. Baffet a insisté sur le côté novateur de la méthode utilisée dans cet article, qui est une révolution pour le monde de la biologie cellulaire. En effet, les auteurs ont mis en place une technique qu’on appelle l’imagerie subcellulaire en direct, et qui permet de visualiser en direct les mouvements d’une protéine dans une cellule vivante.
Ici, les chercheurs ont voulu étudier le déplacement des vésicules portant RAB6 (RAB6+). Ils ont donc créé un transgène, une construction contenant un gène d’intérêt créé « artificiellement » et intégré dans les cellules d’un organisme d’intérêt (ici un embryon de souris) pour étudier son effet. Les auteurs ont construit ce transgène avec la partie codante de RAB6, qu’ils ont fusionnée avec un gène rapporteur : une protéine qui a une fonction d’étiquette et qui permet de visualiser une protéine d’intérêt. Le gène rapporteur utilisé dans ces expériences est la GFP (pour Green Fluorescent Protein). Comme son nom l’indique, c’est un gène qui code pour une protéine qui va émettre une fluorescence verte. Ainsi, la protéine RAB6 apparaîtra verte quand ils regarderont au microscope.
Pour injecter ce transgène dans les cellules aRG, ils font une incision dans le ventre d’une souris femelle gestante. Leur but est ensuite d’injecter le transgène dans le cerveau des embryons, dans le milieu qui entoure les cellules. Ils utilisent pour cela une aiguille qui traverse à la fois l’utérus de la mère et le crâne de l’embryon. Ils placent ensuite des électrodes autour du crâne de l’embryon et font passer un petit courant électrique pour que le transgène rentre directement dans la cellule aRG. Ce processus s’appelle l’électroporation in utero [Figure 6]. Puis, les scientifiques replacent l’utérus dans la cavité abdominale de la mère pour laisser le gène s’exprimer pendant quelques jours. Enfin, les cerveaux des embryons sont récupérés puis coupés en sections assez épaisses (300 µm). Les cellules restent vivantes pendant environ une heure, ce qui permet d’observer les coupes au microscope à très haute résolution.
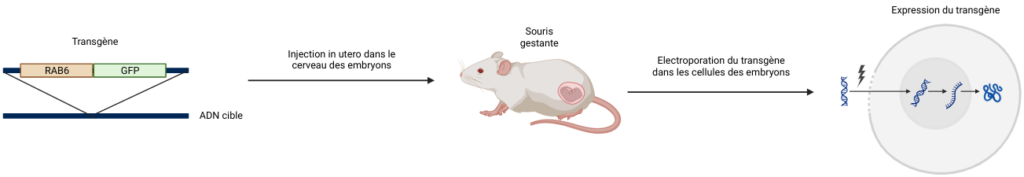
Cette technique permet donc d’étudier la localisation spatio-temporelle d’une protéine, ce qui peut être très utile pour répondre à de nombreuses questions biologiques !
[*] Un progéniteur est un ancêtre fondateur d’une descendance. Dans notre cas, une cellule progénitrice permet de produire d’autres cellules par division/multiplication. Ces dernières cellules peuvent, elles, se différencier en cellules du système nerveux comme les neurones, les astrocytes, les oligodendrocytes ou des cellules gliales.
[**] Une vésicule est une structure sphérique formée d’une membrane biologique et pouvant contenir des molécules qui sont ainsi transportées jusqu’au lieu où elles exerceront leur fonction.
[1] Rakic P., Evolution of the neocortex: a perspective from developmental biology. Nature reviews neuroscience, 2009. DOI : 10.1038/nrn2719. [Review]
[2] Ferent J., et al., Extracellular Control of Radial Glia Proliferation and Scaffolding During Cortical Development and Pathology. Frontiers Cell Dev. Biol., 2020. DOI : 10.3389/fcell.2020.578341. [Review]
[3] Penisson M., et al., Genes and Mechanisms Involved in the Generation and Amplification of Basal Radial Glial Cells. Frontiers Cell. Neurosci., 2019. DOI : 10.3389/fncel.2019.00381. [Review]
[4] Marchadier A., Analyses d’images de tomographie X chez le petit animal : applications aux études de phénotypage ex vivo et in vivo, 2011. [Manuscrit de thèse de doctorat]
Publié le 26/04/2024
 E. Mary/F. Bouvard/G. Maillard/P. Guernalec/Papier-Mâché/CC BY-NC-SA 4.0 2024
E. Mary/F. Bouvard/G. Maillard/P. Guernalec/Papier-Mâché/CC BY-NC-SA 4.0 2024Texte et images, à l’exception des images soumises à droits d’auteurs différents : voir au cas par cas en légende.