Mots clés : Billets de Blog, Bionum
Etiquettes: Apiacées, Charles Darwin, Depressaria radiella, Ecologie Evolutive, Fruits, Fruits sans graines, Graines, Lewis Sturtevant, Panais, pappus, parthénocarpique, Pastinaca sativa

Savoir, c’est comprendre comme la moindre chose est liée au tout – Alain, 1925.
En voilà une bien belle citation ! Elle décrit parfaitement la jeune science qu’est l’écologie, c’est-à- dire l’étude des interactions entre les êtres vivants et leur environnement.
Cette science, déjà sacrément bordélique par sa pluridisciplinarité, finit d’achever nos pauvres cerveaux quand elle s’associe à la biologie évolutive. L’étrange chimère qui en ressort s’appelle « écologie évolutive », et c’est cette discipline que je vais m’évertuer à vous vendre à travers ce billet.

Je l’aime, cette discipline. Aussi bordélique qu’elle soit.
Elle recèle d’énigmes toutes plus complexes les unes que les autres, et dont les solutions vous offrent un sentiment si particulier de travail accompli.
Les avancées scientifiques en écologie évolutive résultent ainsi des efforts de dizaines de générations de biologistes, finissant leurs carrières sans un cheveu sur le caillou, tant ils se les sont arrachés devant les mystères de notre bien-aimée nature.

L’énigme, que dis-je, l’enquête que nous allons mener dans ce billet, c’est celle des fruits sans graines, considérés pendant longtemps comme des aberrations de la nature (le fruit n’ayant, a priori, pas d’autre rôle que celui de protéger et disséminer les graines).
Ainsi, le botaniste Edward Lewis Sturtevant, dans un article publié en 1890 dans la série des Memoirs of the Torrey Botanical Club décrivit la chose comme un exemple d’événement nettement préjudiciable à la conservation et à la distribution des espèces, qui, à moins d’être corrélés à un développement ou à une augmentation d’autres moyens de propagation,[..] devrait entraîner à la destruction de la variété.
Alors, les fruits sans graines ne sont-ils que des erreurs, des bugs de la génétique, ou peut-on trouver des exemples, dans la nature, d’occurrence programmée du phénomène ?
En d’autres termes, peut-on trouver un avantage évolutif au développement de fruits sans graine ?
I/ Les fruits
Mais tout d’abord, à quoi ça sert, un fruit ? Pourquoi les plantes dépensent-elles tant d’énergie pour former ces machins ?
Aussi invraisemblable que cela puisse paraître, leur utilité première n’est pas de finir dans vos yaourts : ils jouent un rôle essentiel dans la reproduction chez les plantes.



Pour disséminer les graines, les plantes ont, au cours de l’évolution, inventées tout un tas de tactiques toutes plus ingénieuses les unes que les autres :

(Bon, techniquement, la fraise est un “faux fruit”, c’est-à-dire un truc qui prend l’apparence d’un fruit mais n’en est pas vraiment un. Le principe est le même : si c’est coloré et sucré, c’est bien pour attirer les animaux !)
Certaines produisent des fruits colorés, sucrés, riches en nutriments, pour attirer les animaux.
D’autres développent de petits parachutes (les pappus) ou de petites ailes sur les fruits, pour favoriser leur transport par le vent.
Bon en vrai les pappus ça ressemble plus à ça..
Enfin, certaines produisent des fruits si léger qu’ils peuvent flotter sur l’eau.

Un fruit est dit « parthénocarpique » quand il ne contient aucune graine.
Par d’habiles mélanges hormonaux, on peut créer artificiellement des variétés de fruits sans graines : cela à un certain intérêt commercial, vous empêchant par exemple de vous casser les dents en mangeant une banane (eh non, les petits trucs noirs à l’intérieur ne sont pas des graines, mais des ovules non fécondées ).
 Les bananes sauvages ont l’air nettement moins appétissantes.
Les bananes sauvages ont l’air nettement moins appétissantes.
La présence naturelle de fruits parthénocarpiques, de manière récurrente chez certaines espèces, est, par contre, plus difficile à expliquer. La formation d’un fruit représente une quantité non négligeable d’énergie gaspillée, et l’unique rôle d’un fruit étant (a priori) la perpétuation de l’espèce, un tel phénomène peut, au mieux, être considéré comme une bizarrerie, au pire, comme un coup de sandale sur le front de Darwin.

Le bougre a pourtant, dans nombre de ses écrits, tenté de répondre à cette question, sans jamais en trouver un réel avantage évolutif.
Il avança notamment qu’il puisse s’agir d’une stérilisation progressive due à la domestication par l’homme, ou la conséquence d’environnements défavorables à la production de graines (en altitude, par exemple), obligeant la plante à se reproduire sans passer par la fécondation (Darwin, 1868).
Les bases ayant été posées, passons à notre enquête…
II. Les protagonistes
Toute bonne enquête commence avec des personnages principaux beaux et charismatiques. Malheureusement, notre premier protagoniste n’est ni beau, ni charismatique, et même si je m’étais évertué à vous le décrire de la manière la plus stylée possible, il ne vous aurait suffi que d’une photo pour décrédibiliser tout mon discours.
Mesdames et Messieurs, voici le panais.
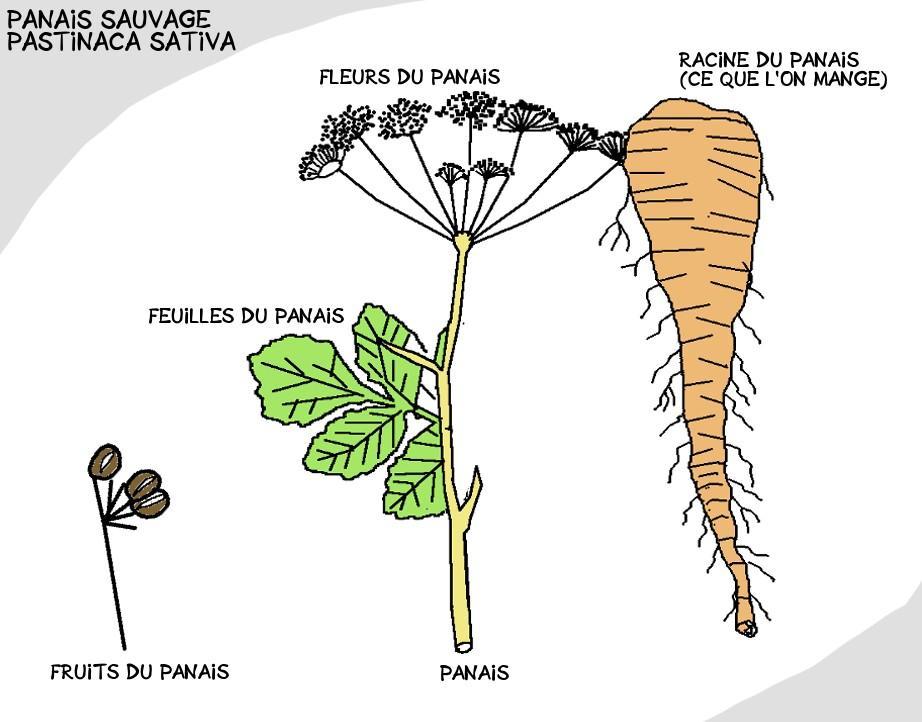
Si ce magnifique légume ressemble autant à une carotte, c’est parce que les deux compères appartiennent à la même grande famille : les Apiacées. Cette confusion fut entretenue durant des siècles et il fallut attendre le XVIIIe siècle pour qu’ils soient officiellement séparés en deux genres bien distincts.
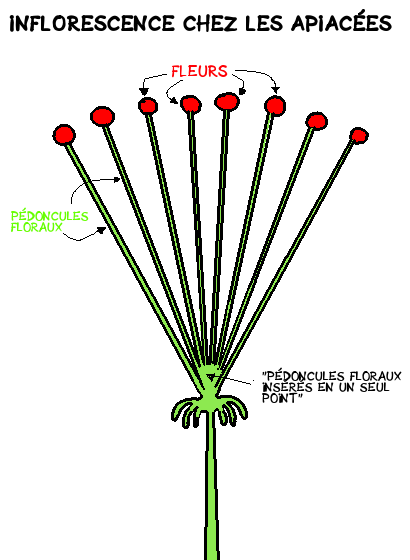
Les Apiacées ont comme caractéristique commune de former, sur un même individu, des groupements de fleurs appelées inflorescences, dont les pédoncules – petites ‘tiges’ portant la fleur- sont insérés en un seul point.
Je pense dorénavant avoir perdu la moitié de mes lecteurs : le panais a le don de rendre inintéressant tout ce que l’on dit, tant tout ce qu’il dégage n’inspire que l’ennui.

J’espère regagner votre attention en vous parlant de son pire ennemi : Celui-Dont-On-Ne-Connait-Pas-Le-Nom. Ce pauvre herbivore de la famille des papillons est tellement peu connu en France que personne n’a daigné le nommer autrement que par son nom latin (Depressaria radiella).
En anglais, ils sont appelés « parsnip webworm », que l’on peut maladroitement traduire en « pyrale des panais », pyrale étant un terme générique désignant certaines espèces de papillons ravageurs.
C’est donc ce malheureux anonyme (au sens étymologique du terme, an-numos : sans nom) qui constituera notre second protagoniste.
Avant de nous pencher sur les habitudes de vie de cet étrange personnage, je me dois de vous détailler sa phylogénie (sa famille), puisque Pierre Kerner, grand amoureux des insectes, sera l’un de mes premiers lecteurs, et il serait malvenu de ne pas le caresser dans le sens de la seta (“poil” chez les insectes – je suis vraiment très rigolo).

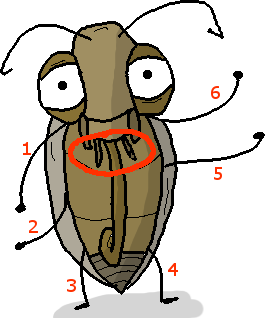
La pyrale est un insecte. Rappelez-vous de mon premier billet : il a six papattes, et des pièces buccales externes.
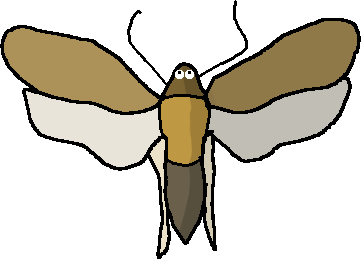
En plus d’être un insecte, il possède de majestueuses ailes, faisant de lui un Ptérygote.
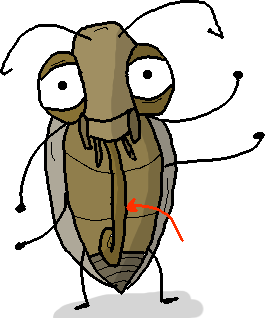
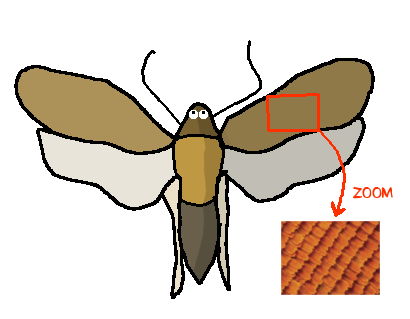
Il est doté d’une trompe, et ses ailes sont composées de pleins de petites écailles : c’est un lépidoptère (donc un papillon).
Enfin, il ne possède pas d’antennes en forme de massue (c’est la caractéristique principale des papillons de jour). C’est donc un papillon de nuit, un hétérocère !

La plupart des œufs de pyrales sont pondus sur les feuilles des panais. Après éclosion, la pyrale, alors sous forme de chenille, va se creuser un petit tunnel de soie dans les bourgeons de la fleur, ou dans les fruits en cours de développement. Elle va s’y nourrir jusqu’à maturité.
Elle formera alors un cocon de caca et de soie et entre en stade de chrysalide, dernière forme avant sa forme finale : l’imago.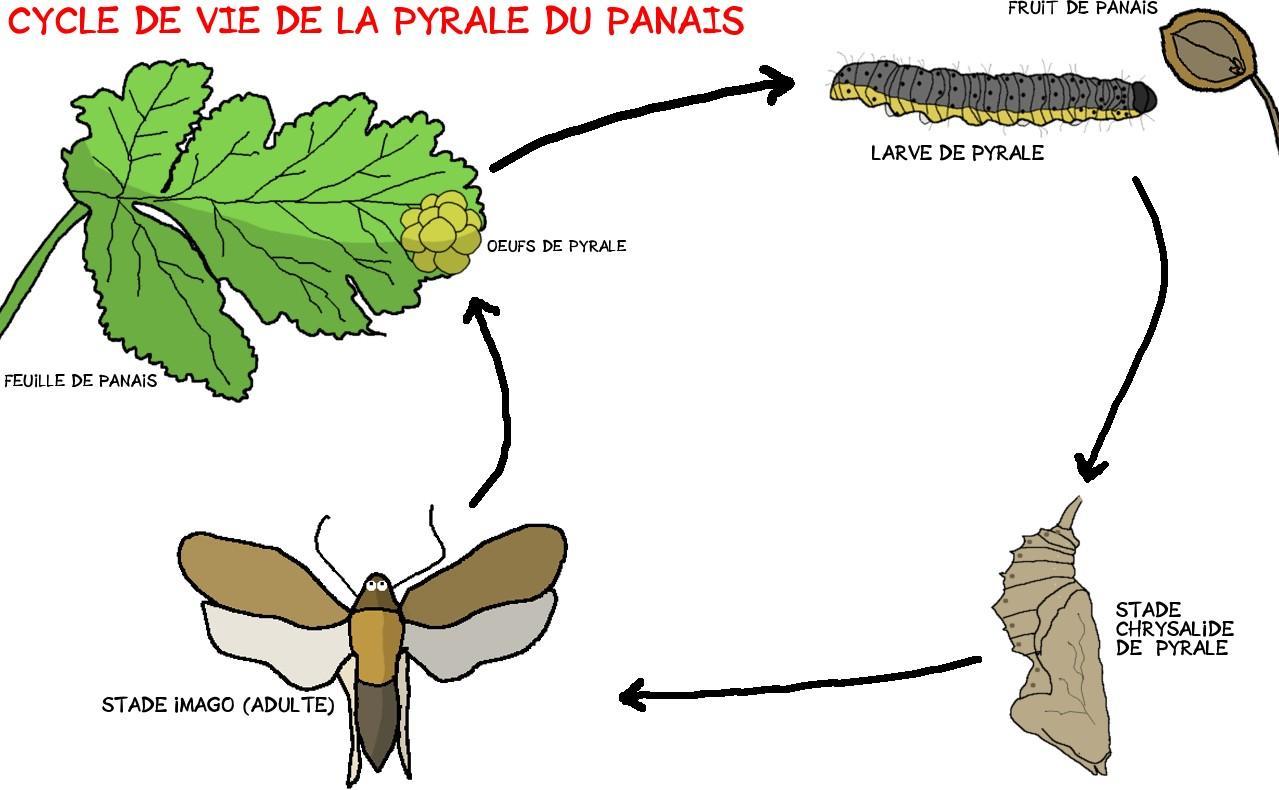
La pyrale est un herbivore (si l’on veut être plus précis, c’est un déprédateur, c’est-à-dire un animal commettant des dégâts sur une plante dans le but de se nourrir). Cela a un certain coût pour le panais, allant au-delà de la simple destruction physique des structures reproductrices : un panais infesté par une seule pyrale produira moins de graines, et sera visité moins fréquemment par les pollinisateurs.

Ces effets secondaires représentent une baisse importante du succès reproducteur du panais, impliquant nécessairement une baisse de ce qu’on appelle, en écologie évolutive, la ‘valeur sélective’, c’est-à-dire la capacité qu’aura un individu, selon ses différentes caractéristiques génétiquement programmées, à se reproduire et ainsi transmettre son patrimoine génétique aux générations suivantes.
Cette valeur sélective sera le principal moteur de la sélection naturelle : un individu ayant un caractère conférant un avantage significatif par rapport à ses congénères aura plus de chance de se reproduire. Il en sera alors de même pour ses descendants, et ainsi de suite : après un nombre important de générations, ses descendants formeront la majorité de la population.
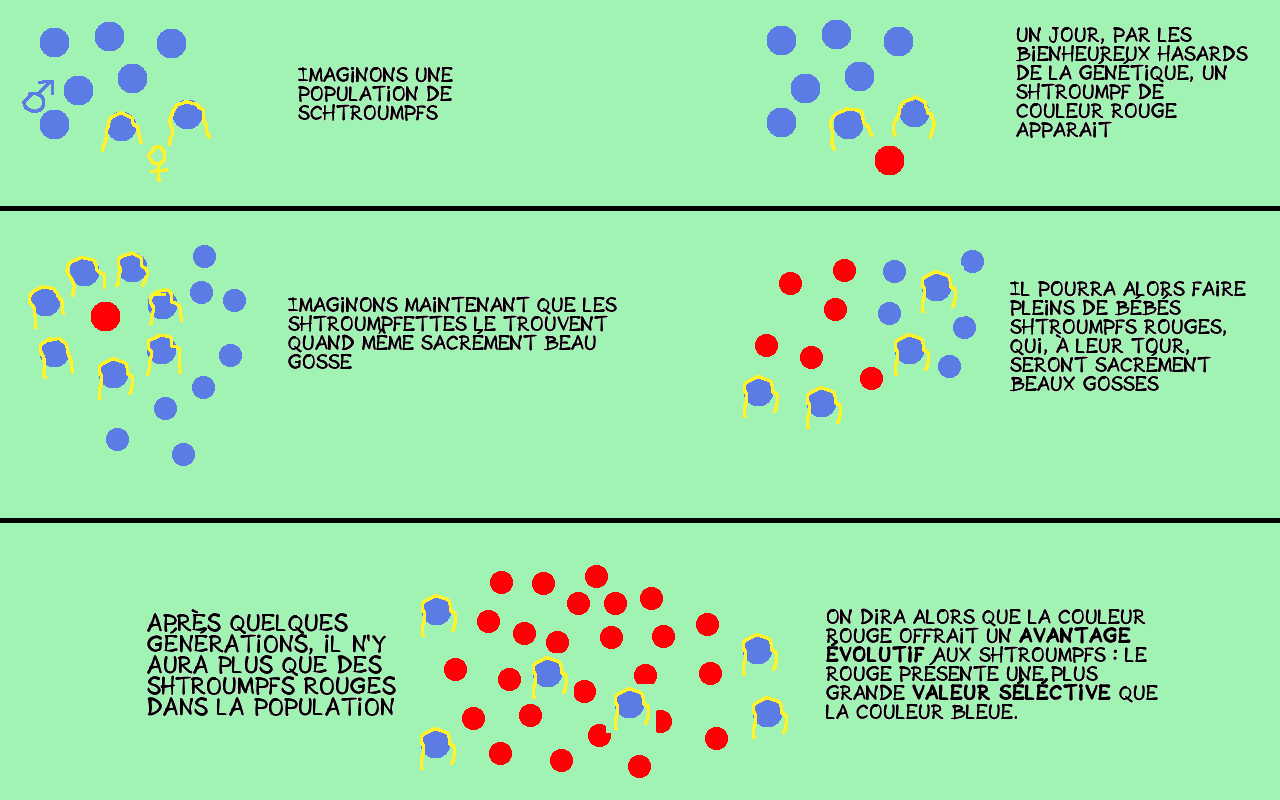
Ces avantages évolutifs peuvent prendre des formes très particulières, voire absurdes, puisque, du point de vue de la nature, toute tactique est bonne pour se reproduire.
Imaginons qu’un individu X, parmi la population humaine, naisse avec un gène particulier lui conférant une patience et une détermination à toutes épreuves, lui permettant, par exemple, d’écrire des billets de blogs durant les grandes vacances, alors qu’il fait beau dehors et qu’il pourrait sortir en ville avec ses amis. Imaginons maintenant que le sexe opposé soit particulièrement attiré par les individus capables d’une telle abnégation. Il va sans dire que l’individu X, ce beau gosse des temps modernes, aura alors un succès reproducteur phénoménal, et, qu’après de nombreuses générations, ce caractère déterminé et patient soit partagé par toute la population humaine.
Chez les panais, la présence de la pyrale implique une baisse du succès reproducteur : cela représente pour les plantes un désavantage évolutif certain. Au cours de l’évolution, seront alors sélectionnés des individus capables de minimiser l’impact de l’herbivorie, voir de l’éviter. Et cela peut prendre, dans la nature, des formes diverses et variées : des épines, des feuilles au goût très amer (et donc peu appétissantes), des toxines, des camouflages, ..
Il vous faut maintenant réaliser que, du point de vue de l’herbivore, un caractère lui permettant de manger plus sera un caractère avantageux : vous avez une idée du joyeux bordel que constitue l’écologie évolutive.

Cette course à l’armement évolutif, que l’on retrouve de manière générale dans les relations proie- prédateur ou parasite-hôte, s’appelle la co-évolution, phénomène que j’ai eu l’occasion de vous évoquer lors de ma participation à la conférence L’Arche de Diderot.
Revenons à nos panais.
La destruction des structures reproductrices ainsi que des fleurs, conduit nécessairement à l’altération des repères d’orientation chimique utilisés par les pollinisateurs, ainsi que la diminution de l’accès aux fleurs en raison de la présence des soies produites par les pyrales.
À cause des pyrales, les panais sont moins visités par les pollinisateurs, il y a ainsi une baisse de la fécondité et donc une baisse du nombre de graines produites. Enfin, une proportion conséquente des graines n’arrive même pas à maturité à cause des pyrales se nourrissant des fruits en cours de développement.
La présence de la chenille représente une véritable catastrophe pour les panais.
Maintenant que je vous ai présenté les différents protagonistes, ainsi que les interactions qu’il peut y avoir entre les deux, il est temps que nous rentrions dans le vif du sujet.
III/ Les éléments de l’enquête
Mon objectif est que vous vous trituriez un peu le crâne en lisant ce billet de blog, je vais donc vous présenter les différents éléments mis en évidence par les chercheurs, sans vous dire toute de suite le fin mot de l’histoire.
Comme dans toute bonne enquête, j’ai glissé des éléments inutiles qui ne vous serviront aucunement pour la résoudre, saurez-vous les retrouver ?
Au cours de l’évolution, le panais a développé des défenses contre les pyrales, et notamment la synthèse de toxines, les furanocoumarins, pouvant potentiellement causer des dommages irréversibles à l’ADN.
Les pyrales sont capables de détecter la quantité de toxine en goûtant la surface du fruit.
Le panais peut se déguster aussi bien cru que cuit, en potage ou en gratin.
En comparant les quantités de toxine entre les fruits avec ou sans graine, on constate que les fruits parthénocarpiques présentent une concentration inférieure en toxine.
Les panais produisent environ 20 % de fruits parthénocarpiques, qui sont de la même taille et de la même forme que les fruits normaux. En revanche, les fruits sans graines présentent une masse inférieure, qui est due à l’absence de l’endosperme, tissu permettant la nutrition de la graine.
Les fruits parthénocarpiques présentent une valeur nutritionnelle bien inférieure à celle des fruits normaux : une pyrale qui consomme un fruit parthénocarpique verra ainsi sa croissance ralentie de près de 60 % par rapport à une autre qui aurait consommé un fruit normal : consommer un fruit parthénocarpique représente ainsi un désavantage certain pour les pyrales.
Certains papillons de nuit ne volent que le jour.
On observe que les chenilles sont plus attirées par les fruits parthénocarpiques : si elles ont le choix, elles iront toujours vers les fruits sans graines. Elles ne commencent à consommer les fruits normaux que lorsqu’il n’y a plus de fruit parthénocarpique.
Vous avez dorénavant tous les éléments de cette enquête. Alors, qu’en pensez-vous ?
IV. Résolution
Cette énigme n’est pas simple, mais elle illustre parfaitement le principe de co-évolution, de course à l’armement évolutif. Car la clé pour démêler ce merdier, c’est de comprendre que tous les éléments cités ne sont pas apparus au même moment au cours de l’évolution, et que ce que nous observons aujourd’hui est le résultat d’une guerre sans relâche, infinie, que se livrent les milliers de générations de pyrales et de panais depuis qu’est apparue cette interaction proie-prédateur.
Dans notre cas, les fruits parthénocarpiques agissent en fait comme un leurre, éloignant les herbivores des fruits normaux, leur permettant ainsi d’arriver à maturité.
Ce leurre, diablement efficace, repose sur les réponses sélectionnées chez les pyrales face aux toxines produites par les panais : les pyrales sont capables de produire une protéine permettant la digestion de la toxine au détriment de leur vitesse de croissance. Elles ont aussi acquis un comportement particulier : en consommant la surface des fruits, les chenilles sont capables d’évaluer la quantité de toxine de ce dernier et ainsi de ne choisir que les fruits avec des doses faibles de toxine.
En produisant des fruits parthénocarpiques, les panais se servent de ce trait comportemental : les pyrales préféreront consommer ces fruits car leur teneur en toxine est plus faible.
Ce leurre est ainsi doublement efficace : il éloigne les pyrales des fruits normaux et ralentis la croissance des pyrales à cause de leur valeur nutritionnelle insuffisante, les exposant plus longtemps aux prédateurs et pathogènes, et donnant plus de temps aux fruits normaux pour se développer.
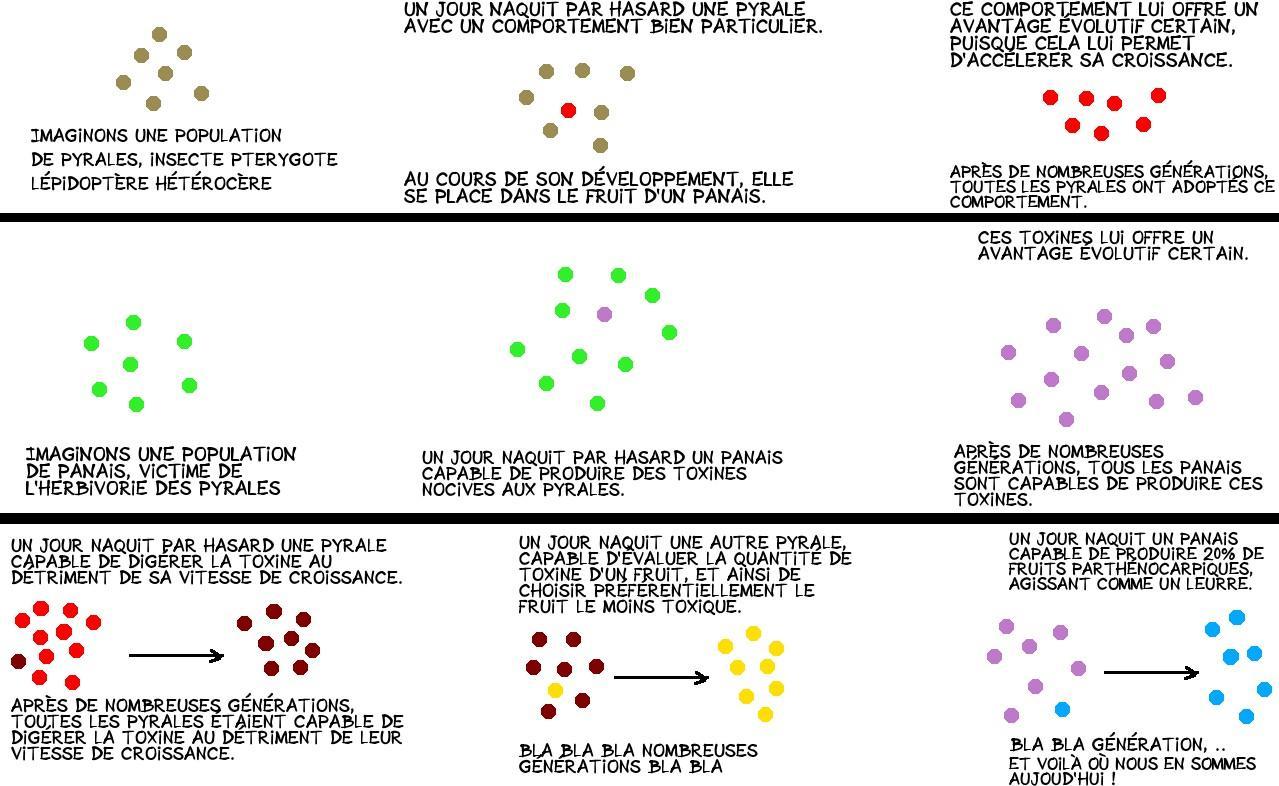
Nous avons ainsi un système de co-évolution entre une proie, le panais, et un prédateur, la pyrale. Au cours de l’évolution, leur interaction a fortement influencé leurs caractères et comportements, l’un pour se développer au dépens du premier, l’autre pour s’en protéger.
Bibliographie (par ordre d’apparition) :
Sturtevant, E. L. (1890) Seedless fruits. Mere. Torr. Bot. Club 1,141-85.
Darwin, C. (1868). The variation of animals and plants under domestication (Vol. 2). O. Judd.
Description du Panais :
Baskin, J. M., & Baskin, C. M. (1979). Studies on the autecology and population biology of the weedy monocarpic perennial, Pastinaca sativa. The Journal of Ecology, 601-610. https://doi.org/10.2307/2259115
Description de la pyrale :
Gorder, N. K. N., & Mertins, J. W. (1984). Life history of the parsnip webworm, Depressaria pastinacella (Lepidoptera: Oecophoridae), in central Iowa. Annals of the Entomological Society of America, 77(5), 568-573. https://doi.org/10.1093/aesa/77.5.568
Description de l’interaction pyrale-panais :
Brittain, W. H., & Gooderham, C. B. (1916). POPULAR AND PRACTICAL ENTOMOLOGY.: An Insect Enemy of the Parsnip. The Canadian Entomologist, 48(2), 37-41.
Impacts de la présence des pyrales sur les panais :
Lohman, D. J., Zangerl, A. R., & Berenbaum, M. R. (1996). Impact of floral herbivory by parsnip webworm (Oecophoridae: Depressaria pastinacella Duponchel) on pollination and fitness of wild parsnip (Apiaceae: Pastinaca sativa L.). American Midland Naturalist, 407-412. https://doi.org/10.2307/2426744
Toxines et effets sur les pyrales :
Berenbaum, M. R., & Zangerl, A. R. (1994). Costs of inducible defense: protein limitation, growth, and detoxification in parsnip webworms. Ecology, 75(8), 2311-2317. https://doi.org/10.2307/1940886
Cianfrogna, J. A., Zangerl, A. R., & Berenbaum, M. R. (2002). Effects of furanocoumarins on feeding behavior of parsnip webworms Depressaria pastinacella. Journal of chemical ecology, 28(7), 1365-1375. https://doi.org/10.1023/A:1016244402019
Hypothèse de la formation de fruits parthénocarpiques :
Zangerl, Arthur R., May R. Berenbaum, et James K. Nitao. « Parthenocarpic Fruits in Wild Parsnip: Decoy Defence against a Specialist Herbivore ». Evolutionary Ecology 5, no 2 (avril 1991): 136-45. https://doi.org/10.1007/BF02270830
Berenbaum, M. R., Zangerl, A. R., & Nitao, J. K. (1986). Constraints on chemical coevolution: wild parsnips and the parsnip webworm. Evolution, 40(6), 1215-1228. https://doi.org/10.1111/j.1558-5646.1986.tb05746.x